| Index to this page |
Could a mutation in one of your liver cells ever be passed on to your children?
No!Why not?

It turns out that the fate of the 16 cells produced by the first 4 mitotic divisions of the zygote is completely predictable. The zygote is, of course, totipotent. That means that it retains the potential to form all the cells — germline and somatic — of the animal. At each of the first four mitotic divisions, one daughter cell (red) loses this totipotentiality and all her daughters will be restricted to forming the somatic tissues of the animal. The second daughter cell — shown in blue in the figure (P1, P2, P3, and P4) retains its totipotentiality. At the 5th mitotic division, both daughter cells remain totipotent and go on, in due course, to form the sperm and eggs. (Any imprinted genes inherited from the mother or father have their imprints removed at this time — link.)
The different fates of the germline (blue) and somatic cells (red) are controlled by certain proteins that are retained in the germline but destroyed in the somatic cells (by ubiquination).
At each generation, the embryo that develops from the zygote not only sets aside some germplasm for the next generation but also produces the cells that will develop into the body, the soma, of the organism.
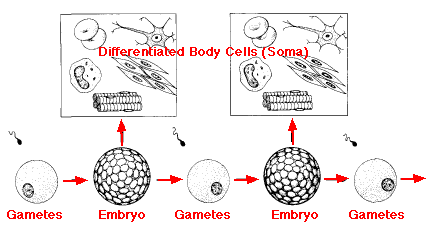
| Today we know that only the germplasm — the gametes and the cells that form them — continue to express high levels of the enzyme telomerase. These cells are able to maintain the length of their chromosomes forever and are immortal. The cells of the somaplasm, in contrast, stop producing telomerase, lose a portion of their chromosome tips at each mitosis, and eventually die. |
In Weismann's view, the somaplasm simply provides the housing for the germplasm, seeing to it that the germplasm is protected, nourished, and conveyed to the germplasm of the opposite sex to create the next generation. The old riddle about which came first, the chicken or the egg, would have been no puzzle to Weismann. In his view, the chicken is simply one egg's device for laying another egg.
Weismann also understood the implications of his theory for aging. Once the opportunity to pass germplasm on has passed, there is no need to maintain the integrity of the somaplasm ("disposable soma"); hence the decline in body function with aging .
In insects in appears to be controlled by the special quality of the cytoplasm deposited by the mother at one end of the egg. [Link to discussion of embryonic development in insects.]
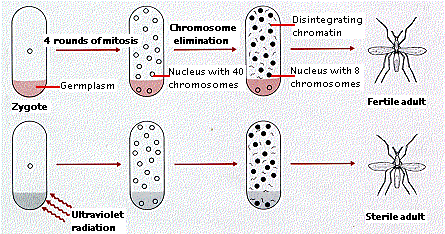
At the 4th mitotic division in the gall midge (an insect) egg, 2 of the 16 nuclei become pinched off in a small amount of cytoplasm at one end of the egg. At the 5th mitosis, these two nuclei divide normally, producing daughter cells with the full complement of chromosomes (2n = 40) of the species.
But not so for the other nuclei. When each of these reaches anaphase, only 8 of their 40 chromosomes (dyads) separate and move to opposite ends of the spindle. The remaining 32 chromosomes stay at the metaphase plate and eventually disintegrate.
The top row of this figure shows normal development in the gall midge. The gametes are descended from the two nuclei, each with a full diploid set of 40 chromosomes, that were partitioned off in a mass of special cytoplasm (here called "germplasm"). The remaining nuclei lose 32 chromosomes before going on to form the rest of the insect body (the somaplasm).
The bottom row shows that destruction of the germplasm causes the nuclei that move there to undergo chromosome elimination also. The animal that develops is sterile but otherwise normal.
In mammals (and birds), the decision to become germ cells is not intrinsic but is the result of cell-to-cell signaling during early embryonic development.
In mice, the setting aside of the future germ cells begins at the start of gastrulation (6 days after fertilization in the mouse). Cells of the future extraembryonic membranes secrete cytokines (including bone morphogenic proteins) which signal cells in the epiblast (the upper layer of cells in the inner cell mass — ICM) which will go on to form ectoderm and mesoderm. Under the influence of other inter-cellular chemical signals, a few of these epiblast cells will develop into primordial germ cells (PGCs).
The PGCs will later migrate into the part of the developing embryo that will go on to form the gonads (ovaries or testes).
All vertebrates and ecdysozoans (e.g. Drosophila) segregate their germline from somatic tissue just as Weismann postulated; that is very early in embryonic development an irreversible process takes place so that somatic cells can never give rise to germline cells and vice-versa.
However, there are some exceptions.
Adult planarians and at least some cnidarians (e.g. Hydra) retain a population of stem cells throughout life that give rise to gametes as well as more somatic tissue.
All sponges also contain a population of adult stem cells which give rise to both somatic cells and germline cells throughout the animal's life.
In plants as well, cells destined to become gametes do arise from somatic tissues. In the flowering plants (angiosperms), for example, certain signals cause meristems that had been making stem tissue to become converted into flower buds which go on to make the gametes.
| see Plant Growth and also a discussion of Flowering |
In microorganisms, all life's functions are embodied in a single cell. (However, some unicellular organisms, like the ciliated protozoan Tetrahymena thermophila, have a complete genome in their micronuclei, which are passed on to the next generation, as well as genes in a macronucleus, which is not. Thus, even here, there is the equivalent of a distinction between germline and soma.)
The significance of mutations is profoundly influenced by the distinction between germline and soma. Mutations that occur in a somatic cell, in the bone marrow or liver for example, may
A germline mutation, in contrast, will be found in roughly half the cells descended from the zygote to which that mutant gamete contributed. If an adult is successfully produced, roughly half of its cells will contain the mutation. Included among these will be the next generation of gametes, so if the owner is able to become a parent, that mutation may pass down to yet another generation.
| Welcome&Next Search |