| Index to this page |
First, Darwin presented a wealth of evidence of evolution. He said that all living things on earth today are the descendants — with modifications — of earlier species.
Second, he proposed a mechanism — natural selection — to explain how evolution takes place.
Evolution involves two interrelated phenomena:Over the course of time, species modify their phenotypes in ways that permit them to succeed in their environment.
This page is devoted to looking at how evolution leads to adaptation.
Over the course of time, the number of species multiplies; that is, a single species can give rise to two or more descendant species. In fact, Darwin maintained that all species are related; that is, any two species on earth today have shared a common ancestor at some point in their history.
The process of speciation is examined in a separate page.| Link to it. |
The forces of natural selection act on phenotypes, but only if there is a change in the genotypes of a population has evolution occurred.
| Population genetics is the study of the genotypes and their changes in entire populations. Link to a discussion. |
| Link to discussion of how polymorphism of MHC alleles protects against parasites. |
In other cases of sexual selection, one phenotype prefers to mate with others of the same phenotype. This is called assortative mating.
All the forces of natural selection outlined above work on individuals. But there is an increasing body of evidence that natural selection can also act on groups. Natural selection that appears to work counter to the benefit of some individuals while enhancing the prospects of their relatives is called kin selection. It is discussed on a separate page. Link to it.
of all the human eggs that are fertilized, only a small percentage will ever reproduce themselves.
The others are eliminated as follows:
A plot of the distribution of the trait in a population often produces a bell-shaped curve like this one that shows the distribution of heights among a group of male secondary-school seniors.
Such a distribution could arise from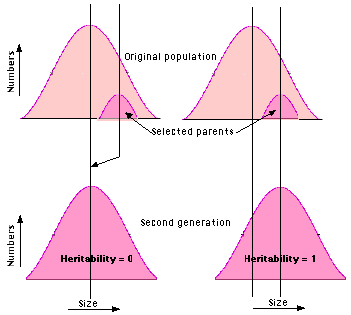
One can sort out the relative contribution of genetic and environmental factors by comparing the range of a trait in the offspring compared with the average value of that trait in their parents.
If the offspring of selected parents occupy the same range as the entire population, environmental factors are working alone. The trait has a zero heritability.
Example: The length of the seeds of a pure strain of beans may vary over several millimeters. However, if extra-large beans are mated, the new crop shows no shift to a larger size. So the heritability of length is zero.
On the other hand, if the offspring of two extra-large mice are just as large as they are, genes are probably at work. The trait is said to have a heritability of 1.
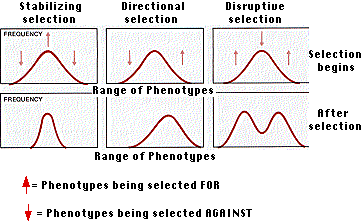 Natural selection often works to weed out individuals at both extremes of a range of phenotypes resulting in the reproductive success of those near the mean. In such cases, the result is to maintain the status quo.
Natural selection often works to weed out individuals at both extremes of a range of phenotypes resulting in the reproductive success of those near the mean. In such cases, the result is to maintain the status quo.
It is not always easy to see why both extremes should be handicapped; perhaps sexual selection or liability to predation is at work. In any case, stabilizing selection is common. In humans, for example, the incidence of infant mortality is higher for very heavy as well as for very light babies.
Since 1973, Peter and Rosemary Grant — aided by a succession of colleagues — have studied Darwin's finches in the Galapagos Islands.
| Link to drawings of Darwin's finches. |
When rainfall, and thus food, are plentiful, the ground finches tend to
 The graph (from P. T. Boag and P. R. Grant in Science 214:82, 1981) shows how the population declined from 1400 to 200 on the island of Daphne Major, a tiny (10-acre = 4 hectares) member of the Galapagos Islands.
The graph (from P. T. Boag and P. R. Grant in Science 214:82, 1981) shows how the population declined from 1400 to 200 on the island of Daphne Major, a tiny (10-acre = 4 hectares) member of the Galapagos Islands.
| Link to map. |
One of the plants to make it through the drought produces seeds in large, tough fruits that are virtually impossible for birds with a beak smaller than 10.5 mm to eat.
Sampling the birds that died as well as those that survived showed that| Beak length (mm) | Beak depth (mm) | |
| Dead birds | 10.68 | 9.42 |
| Survivors | 11.07 | 9.96 |
But did it produce evolution?
The answer turned out to be yes. As the population of G. fortis recovered after the rains returned, the average body size and beak depth of their offspring was greater than before (an increase of 4–5% for beak depth). The bell-shaped curve had been shifted to the right — directional selection.
More recently, the Grants and colleagues at Harvard Medical School have shown that
|
 Many species of moths in the British Isles began to become darker in color in the 19th century.
Many species of moths in the British Isles began to become darker in color in the 19th century. The best-studied example is the peppered moth, Biston betularia. The moth gets its name from the scattered dark markings on its wings and body.
In 1848, a coal-black mutant (now known to result from insertion of a transposon) was found near Manchester, England. Within a century, this black form had increased to 90% of the population in this region.
The moth flies at night and rests by day on tree trunks. In areas far from industrial activity, the trunks of trees are encrusted with lichens. As the photos show, the light form (circled in red) is practically invisible against this background.
In areas where air pollution is severe, the combination of toxic gases and soot has killed the lichens and blackened the trunks. Against such a background, the light form stands out sharply.
The moth is preyed upon by birds that pluck it from its resting place by day. In polluted woods, the dark form has a much better chance of surviving undetected. When the English geneticist H. B. D. Kettlewell (who supplied the photos) released moths of both types in the woods, he observed that birds did, indeed, eat a much higher fraction of the light moths he released than of the dark.Since pollution abatement programs were put in place after World War II, the light form has been making a comeback in the Liverpool and Manchester areas.
An example:
The residues ("tailings") of mines often contain such high concentrations of toxic metals (e.g., copper, lead) that most plants are unable to grow on them. However, some hardy species (e.g. certain grasses) are able to spread from the surrounding uncontaminated soil onto such waste heaps. These plants develop resistance to the toxic metals while their ability to grow on uncontaminated soil decreases. Because grasses are wind pollinated, breeding between the resistant and nonresistant populations goes on. But evidently, disruptive selection is at work. Higher death rates of bothThe evolutionary significance of disruptive selection lies in the possibility that the gene pool may become split into two distinct gene pools. This may be a way in which new species are formed.
The formation of one or more species from a single precursor species is called speciation. It is the topic of a separate page.| Link to Speciation. |
| Welcome&Next Search |
{kind=link}