| Index to this page |
In the embryonic development of a zygote, gradients of mRNAs and proteins, deposited in the egg by the mother as she formed it, give rise to cells of diverse fates despite their identical genomes.
| For a discussion of the evidence that leads to this important conclusion, examine Embryonic Development: Getting Started. |
But is the embryo fully patterned in the fertilized egg? It is difficult to imagine that the relatively simple gradients in the egg could account for all the complex migration and differentiation of cells during embryonic development. And, in fact, the answer is no. However, once these gradients have sent certain cells along a particular path of gene expression, the stage is set for those cells to begin influencing nearby cells to become increasingly diversified.
In other words,Cell-cell interactions could — and probably do — occur in several ways:
She cut out a piece of tissue from the gray crescent region of one newt gastrula and transplanted it into the ventral side of a second newt gastrula. To make it easier to follow the fate of the transplant, she used the embryo of one variety of newt as the donor and a second variety as the recipient.
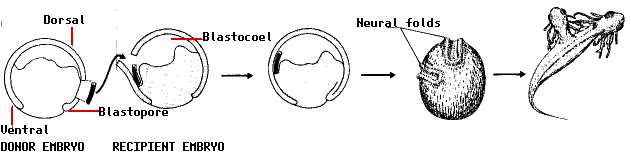
The remarkable results:
But the most remarkable finding of all was that the neural folds were built from recipient cells, not donor cells. In other words, the transplant had altered the fate of the overlying cells (which normally would have ended up forming skin [epidermis] on the side of the animal) so that they produced a second head instead!
Spemann and Mangold used the term induction for the ability of one group of cells to influence the fate of another. And because of the remarkable inductive power of the gray crescent cells, they called this region the organizer.
| Link to another demonstration of induction performed by Spemann. |
Ever since then, vigorous searches have been made to identify the molecules liberated by the organizer that induce overlying cells to become nerve tissue. One candidate after another has been put forward and then found not to be responsible. Part of the problem has been that not until just recently has it become clear that the organizer
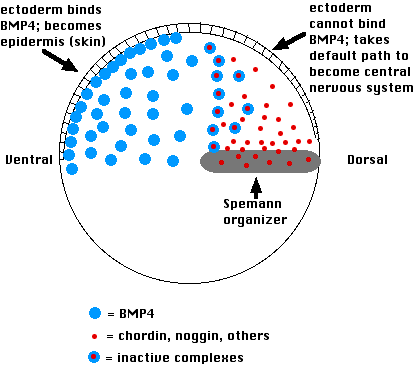
In the Spemann/Mangold experiment, transplanting an organizer to the ventral side provided a second source of chordin. This blocked BMP-4 binding to the overlying ectoderm and thus changed the fate of those cells to forming a second central nervous system rather than skin.
Protein synthesis by the cells of the organizer requires transcription of the relevant genes (e.g., chordin). Expression of organizer genes depends first on Wnt transcription factors. Their messenger RNAs were deposited by the mother in the vegetal pole of the egg. After fertilization and formation of the gray crescent, they
One of the distinguishing features of vertebrates is their tail, which extends out behind the anus.
French researchers have reported (in the 24 July 2003 issue of Nature) their discovery of a tail "organizer", that is, a cluster of cells in the embryo that induces nearby cells to contribute to the formation of the tail.
They worked with the zebrafish, Danio rerio (which also has a head organizer like that of newts).
They removed tiny clusters of cells from the ventral part of the blastula (a region roughly opposite where the Spemann-like organizer forms) and transplanted this into a region of the host embryo that would normally form flank.
The result: a second tail.
Using a fluorescent label, they were able to show that the extra tail was made not only from descendants of the transplanted cells but also from host cells that would normally have made flank.
Three proteins were essential:| Xenopus | Bone Morphogenetic Protein-4 (BMP-4) | blocked by chordin |
|---|---|---|
| and also by noggin | ||
| Drosophila | Decapentaplegic (DPP) | blocked by short gastrulation (SOG) |
| and also by a noggin homolog? |
Although their actions are similar, the distribution of these proteins in Drosophila differs from that in Xenopus (as well as in mammals and other vertebrates).
In Drosophila,The result in Drosophila is that its central nervous system forms on the ventral side of the embryo, not on the dorsal! And, you may remember that one of the distinguishing traits of all arthropods (insects, crustaceans, arachnids) as well as many other invertebrates, such as the annelid worms, is a ventral nerve cord. Chordates, including all vertebrates, have a dorsal (spinal) nerve cord.
| See Embryonic Development: Getting Started |
| see Organizing the Embryo: Segmentation |
| see Embryonic Development: Putting on the finishing touches |
| Welcome&Next Search |